|
| 10. Signals in plants |
| We might think that we are now able to model all possible inflorescences, since we have considered all possible branching types. This is not quite true. |
 One problem is related to the flowering
sequences. For example, in wild carrot, shown in
Plate 21,
flowers first appear at the top of the structure and the flowering zone
progresses basipetally (downwards). As plants develop from the bottom up
and not from the top down, the basipetal flowering sequence is opposite
to the order in which the buds were originally formed.
One problem is related to the flowering
sequences. For example, in wild carrot, shown in
Plate 21,
flowers first appear at the top of the structure and the flowering zone
progresses basipetally (downwards). As plants develop from the bottom up
and not from the top down, the basipetal flowering sequence is opposite
to the order in which the buds were originally formed. |
In order to capture such processes, it is necessary to consider various forms of information flow in a developing plant. The L-system rules outlined in Section 8 represent information transfer by lineage, or from the mother module to the daughter modules. They are called context-free rules in formal language (L-system) terminology. In many plants the interaction between coexisting modules also plays an important role. Such interaction may have the form of a flow of nutrients, water, or hormones between the modules of a developing plant. |
Interactive processes can be captured by context-sensitive L-systems. In the context-sensitive case, a rule is applied to a particular module only if this module has some specific neighbors. |
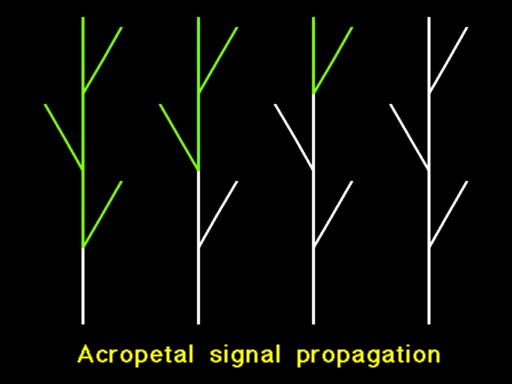
The flow of information in a static structure is of some interest, but even more interesting phenomena occur in growing structures. For example, consider a flower-inducing signal, corresponding to the hypothetical hormone florigen, which is sent from the bottom of the growing plant towards the apices. An apex reached by this signal is transformed into a flowering bud, then a flower, and finally a fruit. |
Depending on the relative growth rates of the main axis and the lateral branches, and on the respective ratios of signal propagation, the flowering zone may progress acropetally (upwards) as shown in Animation 15, or basipetally (downwards) as shown in Animation 16.
Animation 17 and Animation 18 illustrate a basipetal flowering sequence using a schematic and a realistic model of hawkweed (Hieracium umbellatum). Once the apex of the main axis has been transformed into a flowering bud, flowers on the lower branches are formed one after another in a basipetal sequence.
Animation 19 shows a close-up of the development of a single flower from the bud to an open flower to a fruit.
|
Some models may require several signals. This occurs, in particular, when the flowering sequence progresses from the top down, and in some stages the plant is more extensively developed near the top than near the bottom. Mycelis muralis, or wall lettuce, shown schematically in Animation 20, provides an example.
Initially, the apex of the main axis produces a sequence of lateral branches. At this stage, their further development is inhibited. At some point in time, florigen is induced near the base of the plant and propagates upwards. When it reaches the apex of the main axis, the apex is transformed into a bud, and then a flower. Shortly afterwards, another signal is sent from the apex and propagates downwards. This signal lifts the apical dominance, allowing the lateral branches to grow. As this enabling sequence proceeds from the top down, the top part of the structure can be more developed that the bottom part. A third signal (not shown in the animation) introduces florigen into the lateral branches, so that their apices can be eventually transformed into flowers. A three-dimensional rendering of this developmental sequence is shown in Plate 25 and Animation 21.
|